.png)
研究概要
花序の枝分かれパターンを決定する仕組み
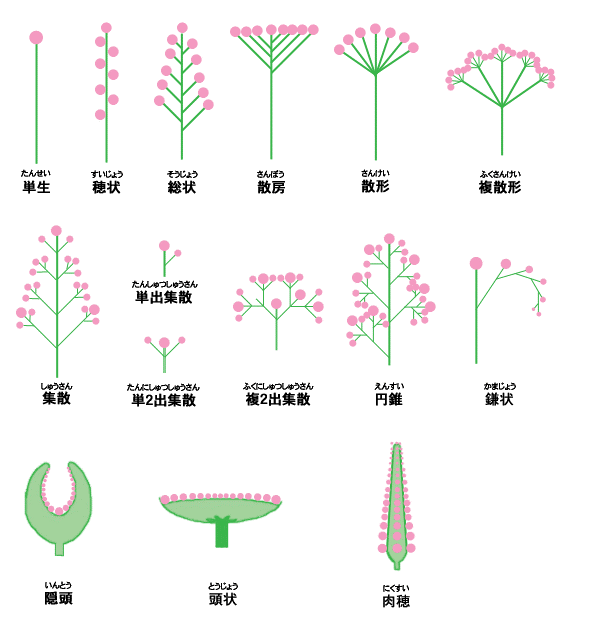
地球上に存在する植物は多種多様です。特に、百花繚乱といわれるように、花序(複数の花がまとまってつく枝)のパターンは多様で美しく、目を奪われます。花序のパターンは植物種に固有であり、総状花序,円錐花序,集散花序,頭状花序等に分類され、植物を分類するうえで重要な形質です (左図)。
このように多様な花序の形態はどのように形成されるのでしょうか。植物の地上部の形態は茎頂メリステムの働きにより決定されます。
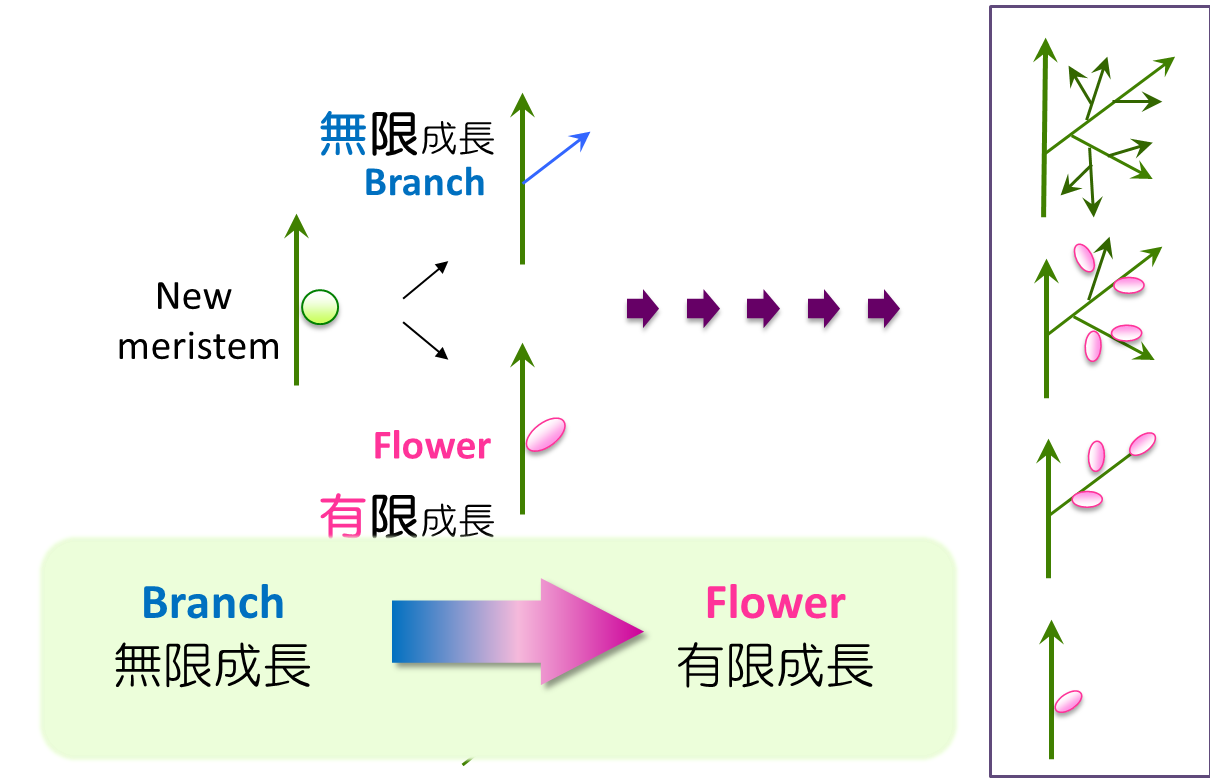
植物の形づくりの基本は「メリステム」です。メリステムに存在する幹細胞の働きにより、器官形成と枝分かれが規則正しく繰り返されます(無限成長性)。季節などの影響により花をつくるように運命づけられたメリステムは花メリステムと呼ばれ、限られた数の花器官を形成した後に幹細胞が失われます(有限成長性)。したがって、メリステムが「花メリステム」としての運命づけられるタイミングとパターンにより花序の形が決まります。また、種子を収穫する作物では、種子数を決定する重要な要因となります。
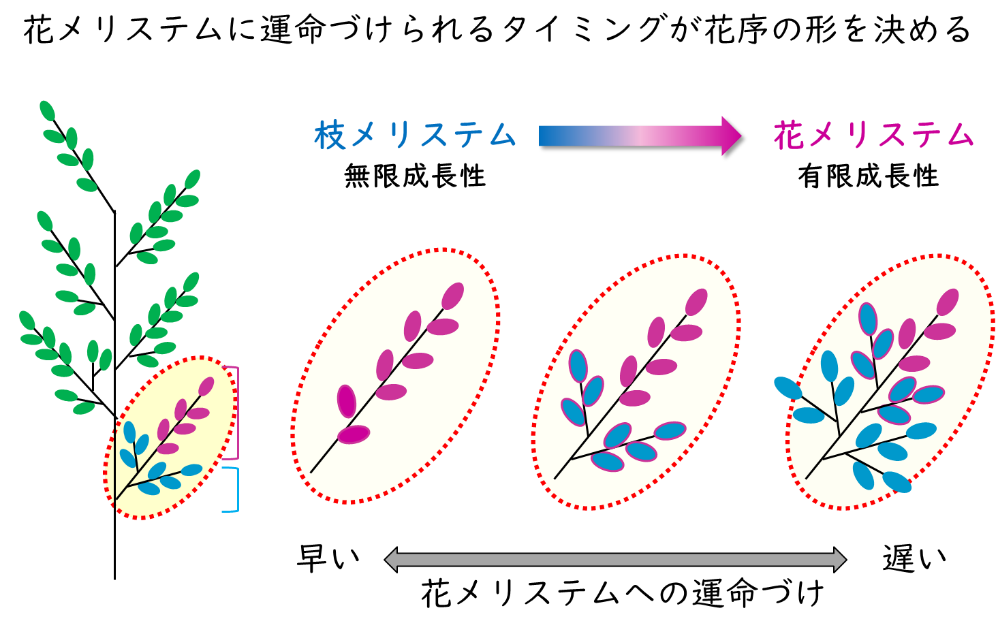
私たちは、イネを対象として花序の形を決めるしくみを研究しています。イネの花序(穂)は茎の先端から一次、二次(まれに三次)の枝分かれが形成され、それぞれの枝分かれに形成されるメリステムは花メリステムになり、花を咲かせ実(コメ)をつけます。花メリステムになるのが遅れるとそのメリステムは枝分かれを続けるの、最終的に枝分かれの多い大きな穂(花の数が多い)を形成します。
花メリステムになるタイミングは、もちろん環境の影響を受けますが、同じ環境で育てる限りは遺伝的に決定されます。すなわち、同じ遺伝子型ならば、同じ条件で育てる限りは同じ穂が形成されます。
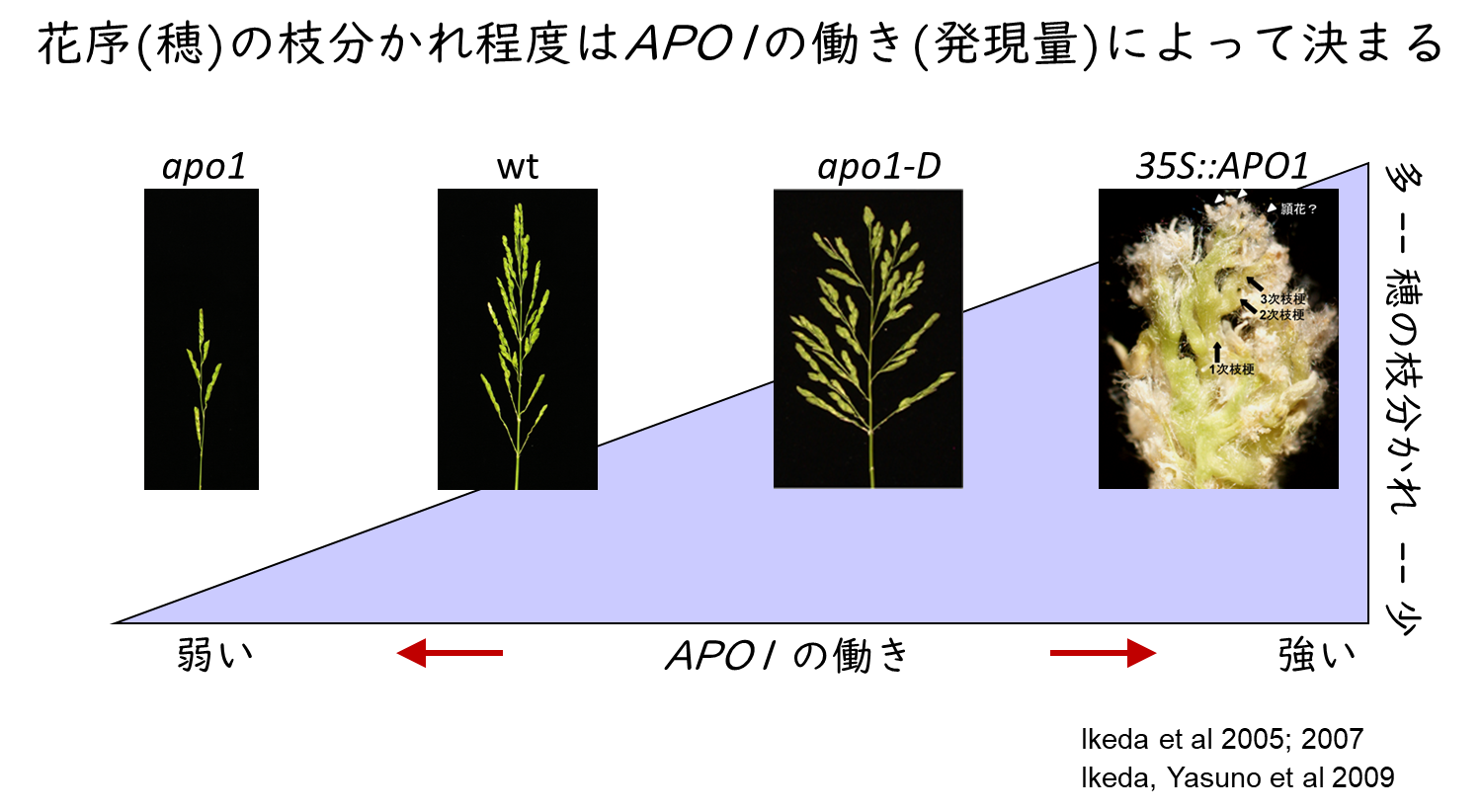
私たちが研究しているTAWAWA1 (TAW1)遺伝子とABERRANT PANICLE ORGANIZATION 1, 2 (APO1, APO2) は、メリステムが花メリステムに運命づけられることを抑える遺伝子です。どちらも、働きが強まると枝分かれが増えます。その結果、花の数が増えます。逆に、機能が弱まると、十分に枝分かれが形成される前にメリステムが花メリステムに運命づけられてしまうため、花の数が減り、小さなメリステムが作られます。すなわち、これら遺伝子の働く程度が花の数(コメの数)を決定します。

私たちが知りたいことは、TAW1やAPO1/APO2がどのように花メリステムへの転換のタイミングをコントロールするのかです。また、これら遺伝子の働く程度がどのような因子により制御されるのかです。