
発表のポイント
- 脳内での情報の受け渡しは、神経細胞同士をつなぐシナプス*1で行われています。学習や記憶にともない、シナプスのサイズが変化し、神経を伝わる信号の強度が制御されます。しかし、シナプス構造の調節メカニズムの全容は不明です。
- シナプスを含む神経細胞の一部を、小脳バーグマングリア細胞*2が断片的に食べる(貪食*3する)ことで、不要な神経接続を弱め、記憶の定着を促進することを示しました。
- 記憶におけるグリア細胞貪食を理解することで、記憶力促進や認知症治療などに役立つ可能性が期待されます。
概要
東北大学大学院生命科学研究科の森澤陽介研究員(研究時、日本学術振興会特別研究員PD)、松井広教授(大学院医学系研究科兼任)らのグループは、小脳バーグマングリア細胞が神経細胞の一部を貪食していることを、新規開発・遺伝子改変マウスを用いた蛍光タンパク質の追跡法*4、三次元電子顕微鏡解析法*5で示しました。さらに、小脳依存性運動学習*6にともない、バーグマングリア細胞によるプルキンエ細胞のシナプス構造の貪食が亢進しました。この貪食作用を薬理学的に阻害すると、シナプス構造の変化が抑制され、学習の効果が低下することが明らかになりました。本研究により、グリア細胞の活躍次第で、記憶の定着のしやすさが左右される可能性が示されました。
本研究成果は、2022年11月1日付でNature Neuroscience誌にAccepted Articlesとして掲載されました。
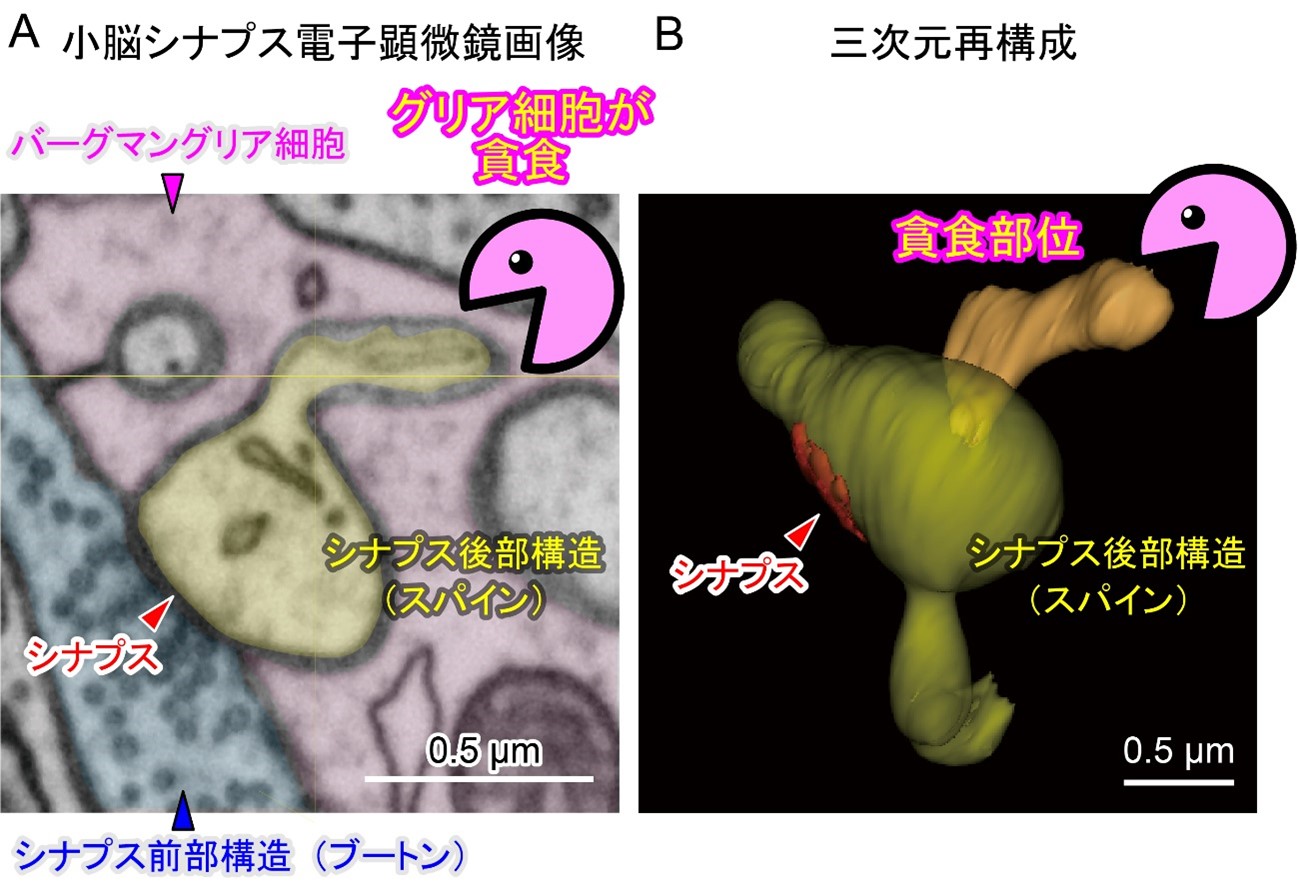
(B) 化学固定された脳組織ブロックをイオン束(Focused Ion Beam)によって薄く削って、残った組織ブロックの表面を走査型電子顕微鏡(Serial Electron Microscope)で観察し、再び、FIBで削って観察することを繰り返すことで、脳組織の精細な三次元像を再構築することが可能です。今回、最新鋭のFIB-SEM技術を使って貪食部位を観察したところ、通常のスパイン形状から明らかに飛び出してグリアに貪食されつつある構造が浮かび上がってきました。
*1 シナプス:
興奮性神経細胞の出力末端のシナプス前部には、グルタミン酸の詰まったシナプス小胞が集積していて、神経細胞が興奮すると、このシナプス小胞の中身が細胞外空間に放出されます(開口放出)。細胞外に放出されたグルタミン酸は、シナプス後部まで拡散し、グルタミン酸に特異的に反応する受容体に結合し、細胞内に陽イオンが流入することで、この神経細胞は興奮します。シナプス伝達の効率が増強されたり減弱されたりすることで短期的な記憶は成立し、シナプスが構造的に変化することで長期的な記憶が定着すると考えられています。小脳においては、シナプス伝達効率が減弱し、シナプス構造が小さくなったり切断されたりすることで、運動学習記憶が形作られると考えられています。
*2 グリア細胞:
脳内の細胞は、神経細胞とグリア細胞に分類されます。神経細胞は活動電位で情報を表現し、神経細胞同士をシナプス結合でつなぐネットワークで脳内情報処理が進むと考えられています。グリア細胞は、神経細胞の隙間を埋めて、神経細胞への栄養供給をする存在だと考えられてきましたが、近年、グリア細胞も特有の情報表現をしていて、神経細胞の担う情報処理に影響を与えることが認識されるようになってきました。
グリア細胞は、大きく分けて、アストロサイト、ミクログリア、オリゴデンドロサイトに分類されます。脳内において情報処理にかかわるのは神経細胞ですが、アストロサイトによる情報処理の修飾やシナプス可塑性のメタ制御等に近年注目が集まっています。今回、神経組織の微細構造が変化し、学習や記憶が神経回路に刻まれて定着する過程において、アストロサイトの貪食作用が関与することが示唆されました。なお、今回、特に注目したのは小脳バーグマングリア細胞についてです。この細胞は、アストロサイトの一種に分類されますが、他のどのグリア細胞とも異なる特徴が多くあるため、独立した細胞種としてみなす場合もあります。バーグマングリア細胞は、特に、小脳依存性の運動学習との関わりが強く示されてきました。
*3 貪食(どんしょく):
細胞が他の細胞の一部や外来の異物を取り囲んで「食べて」しまうように見える現象のことをphagocytosis(貪食)と呼びます。実際に、食べられた細胞の断片は、リソソームという細胞内小器官に取り込まれ、その内容が分解(消化)されます。脳内では、グリア細胞のうちの免疫担当のミクログリアによる脳組織の貪食作用は良く知られていました。また、シナプス部位や血管等の周囲を占め、脳内でのエネルギーの受け渡し等の代謝機能に主な役割をもつアストロサイトも、近年、神経組織を貪食することが示されています。ただ、このようなアストロサイトによる貪食は、発生発達のごく初期の過程か、脳梗塞等によって脳組織の一部が壊れた時にしか生じないと考えられてきました。今回、健常時の脳でも、学習等の刺激によって、アストロサイトによる貪食が亢進することが示されました。
*4 遺伝子改変蛍光タンパク質の追跡法:
今回、神経細胞特異的に蛍光タンパク質を遺伝子発現させて、この蛍光タンパク質の場所を追跡し、グリア細胞に取り込まれたかどうかを調べる方法を用いました。これまで、同様のストラテジーで、神経細胞に緑色蛍光タンパク質(GFP)を発現させて、追跡を試みた例はありますが、貪食された細胞片はグリア細胞内のリソソームに運ばれてしまいます。GFPは早い段階でリソソームによって分解されてしまいます。したがって、貪食された神経組織をGFPで標識して追跡することはできませんでした。一方、今回、リソソームによる分解作用を受けにくい赤色蛍光タンパク質(pHRed)を神経細胞に発現させたところ、グリア細胞内に取り込まれてもなお、pHRedで標識された神経組織を長期的に追跡することが可能になりました。
*5 三次元電子顕微鏡解析法:
グリア細胞には極めて薄いシート状の微細構造があり、光学顕微鏡の解像度を下回るため、グリア細胞が神経組織片を完全に包囲しているのか、それとも、神経細胞とグリア細胞とが、ただ隣接しているだけなのかを区別することは困難でした。また、電子顕微鏡を用いて、超薄切片1枚の画像を調べて、たとえ、神経組織片がグリア細胞に包囲されているように見えたとしても、どこかで神経組織片が神経細胞の本体とつながっている可能性は否定できませんでした。今回、ブロック状に固めた脳組織の表面をFocused Ion Beam(FIB)で薄く削って、ブロック組織の表面を走査型電子顕微鏡(Serial Electron Microscope; SEM)で観察する方法を繰り返すことで、脳組織の三次元像を高解像度で再構築する方法を用いました。この方法によって、神経組織片がグリア細胞によって完全包囲されつつある過程も観察することができ、貪食が完成する前から、神経細胞の微細構造において変性が始まっていることも示唆されました。
*6 小脳依存性運動学習:
各種筋肉の働きを調節して、効率的な運動機能が成り立つような適応学習は、小脳において成立すると考えられています。特に、目の前の画像が左右に触れるのを追跡する不随意の眼球運動の学習(Horizontal Optokinetic Response; HOKR)が良く調べられています。繰り返し、画像を提示することで、次第に眼球運動の振幅が大きくなり、網膜上の画像が安定化すると考えらえられています。この運動学習は、小脳の中でもflocculusという小領域が担当しています。今回、HOKR学習によって、flocculusでのバーグマングリア細胞による神経組織の貪食が亢進し、グリア貪食を薬理学的に阻害すると学習の一部が阻害されることが示されました。
DOI: https://doi.org/10.1038/s41593-022-01184-5